Перспективы отбора - Страница 6
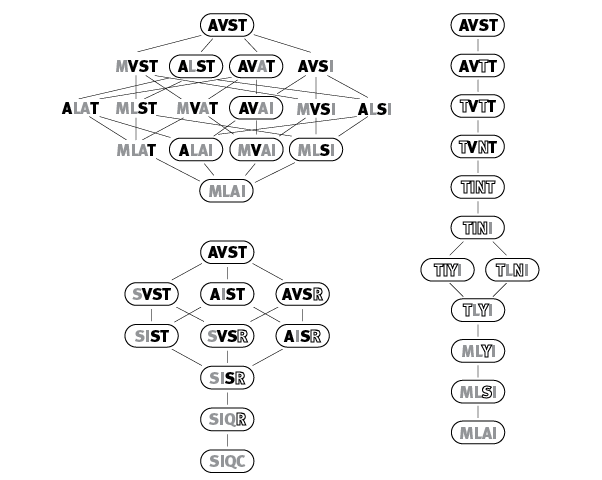
рис. 2.1. Кратчайшие разрешенные эволюционные пути от AVST к последовательностям MLAI и SIQC. Функциональные (разрешенные) последовательности обведены рамками. Контурными буквами обозначены аминокислоты, отсутствующие в начальной и конечной последовательностях. Два левых рисунка в качестве единичного эволюционного события предполагают замену аминокислоты в белке, на правом рисунке таким событием считается замена одного нуклеотида в ДНК. В последнем случае путь получается длиннее из-за структуры генетического кода: от каждой аминокислоты путем замены одного нуклеотида можно прийти не ко всем, а лишь к некоторым из остальных 19 аминокислот. По рисунку из Podgornaia, Laub, 2015.
Во многих случаях все кратчайшие пути оказываются запрещенными. Тогда маршрут из одной точки пространства последовательностей в другую (например, путь от AVST к SIQC, показанный на рис. 2.1 слева внизу) оказывается длиннее, чем кратчайший из потенциально возможных (кратчайший путь предполагает три промежуточных состояния, а на рисунке их четыре).
Ну а если учесть, что реальные единичные мутации обычно представляют собой замену одного нуклеотида в ДНК (а не триплета на любой другой триплет), то проходимость ландшафта окажется еще ниже. На рис. 2.1 справа показан кратчайший разрешенный путь от AVST к MLAI с учетом этого обстоятельства. Теперь он состоит не из четырех, а из целых десяти шагов. Это объясняется структурой генетического кода, которая позволяет путем единичных нуклеотидных замен переходить от данной аминокислоты только к некоторым, но не к любым другим аминокислотам. Например, фенилаланин кодируется триплетами UUU и UUC. Изменив в любом из этих триплетов только одну букву, можно заменить в белковой молекуле фенилаланин на лейцин, изолейцин, валин, серин, тирозин или цистеин. Для перехода к любой из оставшихся 13 аминокислот потребуется не одна, а две или три нуклеотидных замены. Таким образом, эпистаз и структура генетического кода совместными усилиями делают ландшафт приспособленности труднопро- ходимым.
В целом топография изученного плато ландшафта приспособленности оказалась довольно замысловатой. Это плато мало похоже на гладкое, легкопроходимое поле. Оно подразделяется на несколько областей, внутри каждой из которых эволюционирующий белок может передвигаться относительно свободно, однако переход в соседнюю область возможен лишь по немногочисленным тропкам. Некоторые разрешенные последовательности вообще оказываются недосягаемыми из обжитой реальными бактериями области пространства последовательностей. По-видимому, эволюция белок-белкового интерфейса действительно настолько затруднена эпистазом и структурой генетического кода, что эволюция просто не успела разведать многие потенциально доступные области пространства последовательностей.
Данное исследование кому-то может показаться слишком скучным и специальным. Но только не биологам, привыкшим считать “пространство последовательностей” и “ландшафт приспособленности” абстракциями, полезными для теоретических построений, но недосягаемыми для реального изучения. Больше всего впечатляет сам факт, что стало в принципе возможным прямое экспериментальное картирование отдельных областей пространства последовательностей. Хотя, конечно, речь пока идет лишь о четырех аминокислотных позициях, то есть о переборе 204 вариантов. Полное картирование пространства всех возможных белков навсегда останется неразрешимой задачей.
Исследование № 3
“Эволюция умнее, чем ты”: рождение экологического разнообразия
Эволюция не останавливается, организмы приспосабливаются к среде обитания все лучше и лучше даже при неизменных условиях. Но этого мало: даже самая простая среда с точки зрения эволюционирующих в ней организмов оказывается весьма сложной, предоставляющей много альтернативных возможностей. Какой из них следует воспользоваться? Это уж как получится. Одни особи могут повышать приспособленность, подстраиваясь под одни факторы среды, другие – под иные. При этом обе группы, меняясь, неизбежно будут менять и среду обитания друг для друга, и к этим изменениям тоже придется приспосабливаться. В итоге изначально однородная популяция может разделиться на две взаимозависимые, нуждающиеся друг в друге части. Возможно ли такое наблюдать? Оказывается, да. И это еще один замечательный, вполне логичный, хотя и непредвиденный результат долгосрочного эксперимента Ленски, обнародованный в 2017 году. Здесь речь идет о 60 000 поколений. В ходе исследования выяснилось, что за это время как минимум в девяти популяциях из двенадцати произошла экологическая дивергенция: исходно одинаковые бактерии разделились на экологические разновидности. Эти разновидности взаимодействуют друг с другом, сосуществуя вполне по-соседски. Внутри каждой разновидности эволюция продолжается своим ходом, причем дальнейшие изменения направляются как предшествующей эволюционной историей, так и меняющимся экологическим окружением. Таким образом, эволюция перехитрила исследователей, надеявшихся изучить действие мутаций и отбора в “предельно простой” искусственной системе.

Эксперимент Ленски изначально был спланирован так, чтобы свести к минимуму все “осложняющие обстоятельства”: изменения среды, генетический обмен, экологические взаимодействия между организмами. Ученые хотели получить в чистом виде самый главный эволюционный процесс – адаптацию к среде на основе мутаций и отбора. Однако, как метко заметил биохимик Лесли Орджел, “эволюция умнее, чем ты”. Он имел в виду, что исследователям, утверждающим, будто эволюция на что-то не способна, скорее всего, просто не хватает воображения. Как выясняется, эволюция не боится сложностей и “в чистом виде” ничего не демонстрирует, порождая, вопреки чаяниям ученых, куда более замысловатые результаты, чем от нее ждут. В подопытных популяциях Ленски, существующих, казалось бы, в самых простых условиях, какие только можно придумать, стали сами собой зарождаться экологические взаимодействия, основанные на диверсификации (разделении) ниш. А это, в свою очередь, заставляет бактерий заново приспосабливаться к меняющейся биотической обстановке (Good et al., 2017).
На этот раз Ленски и его коллеги провели генетический анализ всей замороженной “ископаемой летописи” эксперимента, накопившейся за 60 000 бактериальных поколений и насчитывающей около 1440 проб (по 120 проб на каждую из двенадцати популяций). Для каждой пробы был проведен метагеномный анализ с 50-кратным покрытием. Это значит, что из пробы выделяли ДНК и секвенировали случайные фрагменты геномов до тех пор, пока каждый участок генома кишечной палочки не оказался “прочтен” в среднем 50 раз. Этого оказалось достаточно, чтобы идентифицировать все новые мутации, которые возникали в подопытных популяциях и достигали частоты не менее 10 % (то есть встречались как минимум у каждой десятой бактерии) хотя бы в двух пробах. Мутации, не получившие столь широкого распространения, не учитывались, потому что их трудно отличить от случайных ошибок секвенирования. В итоге получилась детальная реконструкция эволюционного процесса в двенадцати популяциях.
Выводы о том, что рост приспособленности замедлился, но не прекратился, подтвердились (см. Исследование № 1). Темп накопления новых мутаций остался высоким.
Главное же открытие состоит вот в чем. Динамика накопления мутаций не вписывается в простейшую модель, согласно которой эволюция монокультуры бесполых организмов в стабильных условиях сводится к последовательной фиксации отбором вновь возникающих полезных мутаций. Эта модель не может объяснить наблюдаемую картину даже с учетом таких осложняющих обстоятельств, как генетический автостоп и клональная интерференция, о которых мы говорили выше.