Основы нейрофизиологии - Страница 7
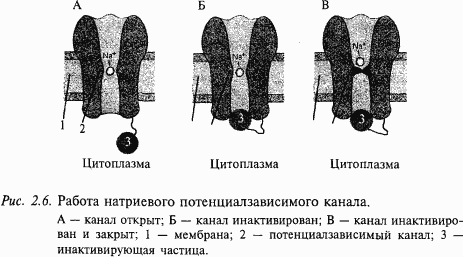
Все изложенное выше можно формально описать следующим образом. В покое клетка ведет себя как «калиевый электрод», а при возбуждении – как «натриевый электрод». Однако после того как потенциал на мембране достигнет своего максимального значения +55 мВ, натриевый ионный канал со стороны, обращенной в цитоплазму, закупоривается специальной белковой молекулой. Это так называемая «натриевая инактивация» (см. рис. 2.6); она наступает примерно через 0,5–1 мс и не зависит от потенциала на мембране. Мембрана становится непроницаемой для натриевых ионов. Для того чтобы потенциал мембраны вернулся к исходному состоянию-состоянию покоя, необходимо, чтобы из клетки выходил ток положительных частиц. Такими частицами в нейронах являются ионы калия. Они начинают выходить через открытые калиевые каналы. Вспомните, что в клетке в состоянии покоя накапливаются ионы калия, поэтому при открывании калиевых каналов эти ионы покидают нейрон, возвращая мембранный потенциал к исходному уровню (уровню покоя). В результате этих процессов мембрана нейрона возвращается к состоянию покоя (–70 мВ) и нейрон готовится к следующему акту возбуждения.
Таким образом, выражением возбуждения нейрона является генерация на мембране нейрона потенциала действия. Его длительность в нервных клетках составляет величину около 1/1000 с (1 мс). Описанная последовательность событий приведена на рис. 2.7.

Подобные потенциалы действия могут возникать и в других клетках, назначение которых – возбуждаться и передавать это возбуждение другим клеткам. Например, сердечная мышца имеет в своем составе специальные мышечные волокна, обеспечивающие бесперебойную работу сердца в автоматическом режиме. В этих клетках также генерируются потенциалы действия (рис. 2.8). Однако они имеют затянутую, почти плоскую вершину, и длительность такого потенциала действия может затянуться до нескольких сот миллисекунд (сравните с 1 мс у нейрона). Такой характер потенциала действия мышечной клетки сердца физиологически оправдан, так как возбуждение сердечной мышцы должно быть длительным, чтобы кровь успела покинуть желудочек. С чем же связан такой затянутый потенциал действия у этого типа клетки? Оказалось, в мембране этих клеток натриевые ионные каналы не так быстро закрываются, как в нейронах, т.е. натриевая инактивация затянута.

Как ясно из этого описания, возбуждение (потенциал действия) нейрона сменяется так называемым «покоем». Однако никакого покоя в этот период нет. Как уже указывалось выше, в мембране есть еще и насосные каналы, количество которых примерно в 10 раз больше ионных, и они постоянно работают, откачивая из цитоплазмы излишек ионов натрия и закачивая туда недостающие ионы калия. Благодаря неустанной работе этих каналов нейрон всегда готов к возбуждению.
Описанный выше механизм возбуждения клетки (конечно, далеко не все клетки нашего организма способны возбуждаться) в основных чертах одинаков не только в нейронах и мышечных клетках человека, но и в аналогичных клетках других организмов. Например, в нейронах моллюсков, червей, крыс и обезьян при возбуждении происходят описанные выше последовательности событий. Более того, конструкция мембран, включая каналы, также примерно одинакова у всех организмов Земли.
Как уже указывалось, каналы представляют собой белковые молекулы, «прошивающие» мембрану (одна часть молекулы находится в цитоплазме, а другая во внеклеточной среде). Интересно, что эти белковые молекулы, образующие ионный или насосный каналы, не вечны, а постоянно заменяются на новые (примерно каждые несколько часов). Все это свидетельствует об очень большой динамичности структуры нейрона.
Нейрон способен к возбуждению, которое состоит в том, что мембрана нейрона в состоянии покоя имеет потенциал порядка –70 мВ (отрицательность в цитоплазме), а в состоянии возбуждения приобретает потенциал +55 мВ. Таким образом, абсолютная величина потенциала действия – около 125 мВ. Длительность потенциала действия нейрона составляет всего около 1 мс (1/1000 с).
Далее это возбуждение (потенциал действия) должно передаться другому нейрону или какой-то другой клетке, например мышечной, железистой и др.
ПРОВЕДЕНИЕ ВОЗБУЖДЕНИЯ
Возбуждение в виде потенциала действия покидает тело нейрона по его отростку, который называется аксоном. Аксоны отдельных нейронов обычно объединяются в пучки – нервы, а сами аксоны в этих пучках называются нервными волокнами. Природа позаботилась, чтобы волокна максимально хорошо справлялись с функцией проведения возбуждения в виде потенциалов действия. Для этой цели отдельные нервные волокна (аксоны отдельных нейронов) имеют специальные чехлы, выполненные из хорошего электрического изолятора (см. рис. 2.3). Чехол прерывается примерно через каждые 0,5–1,5 мм; это связано с тем, что отдельные участки чехла образуются в результате того, что специальные клетки в очень ранний период развития организма (в основном еще до рождения) обволакивают небольшие участки аксона. На рис. 2.9 показано, как это происходит. В периферических нервах миелин образуется клетками, которые получили название шванновских, а в головном мозге это происходит за счет клеток олигодендроглии.

Этот процесс называется миелинизацией, так как в результате образуется чехол из вещества миелина, примерно на 2/3 состоящего из жира и являющегося хорошим электрическим изолятором. Исследователи придают очень большое значение процессу миелинизации в развитии мозга.
Известно, что у новорожденного ребенка миелинизировано примерно 2/3 волокон головного мозга. Примерно к 12 годам завершается следующий этап миелинизации. Это соответствует тому, что у ребенка уже формируется функция внимания, он достаточно хорошо владеет собой. Вместе с тем полностью процесс миелинизации заканчивается только при завершении полового созревания. Таким образом, процесс миелинизации является показателем созревания ряда психических функций. В то же время известны заболевания нервной системы человека, которые связаны с демиелинизацией нервных волокон, что сопровождается тяжелыми страданиями. К самым известным относится рассеянный склероз. Это заболевание развивается незаметно и очень медленно, последствием является паралич движения.
Почему же так важна миелинизация нервных волокон? Оказывается, миелинизированные волокна в сотни раз быстрее проводят возбуждение, чем немиелинизированные, т.е. нейронные сети нашего мозга могут работать с большей скоростью, а значит, более эффективно. Поэтому не миелинизируются в нашем организме только самые тонкие волокна (менее 1 мкм в диаметре), которые проводят возбуждение к медленно работающим органам кишечнику, мочевому пузырю и др. Как правило, не миелинизируются волокна, проводящие информацию о боли и температуре.

Как происходит распространение возбуждения по нервному волокну? Вначале разберем случай немиелинизированного нервного волокна. На рис. 2.10 показана схема нервного волокна. Возбужденный участок аксона характеризуется тем, что мембрана, обращенная к аксоплазме, заряжается положительно относительно экстраклеточной среды. Невозбужденные (покоящиеся) участки мембраны волокна отрицательны внутри. Между возбужденным и невозбужденным участками мембраны возникает разность потенциалов и начинает протекать ток. На рисунке это отражено линиями тока, пересекающими мембрану со стороны аксоплазмы, – выходящий ток, который деполяризует соседний невозбужденный участок волокна. Возбуждение движется по волокну только в одном направлении (показано стрелкой) и не может пойти в другую сторону, так как после возбуждения участка волокна в нем наступает рефрактерность – зона невозбудимости. Нам уже известно, что деполяризация приводит к открыванию потенциалзависимых натриевых каналов и в соседнем участке мембраны развивается потенциал действия. Затем натриевый канал инактивируется и закрывается, что и приводит к зоне невозбудимости волокна. Эта последовательность событий повторяется для каждого соседнего участка волокна. На каждое такое возбуждение тратится определенное время. Специальные исследования показали, что скорость проведения возбуждения немиелинизированных волокон пропорциональна их диаметру: чем больше диаметр, тем выше скорость движения импульсов. Например, немиелинизированные волокна, проводящие возбуждение со скоростью 100–120 м/с, должны иметь диаметр около 1000 мкм (1 мм).