Генетика этики и эстетики - Страница 13
От австралопитеков и питекантропов раннего палеолита нас отделяет 500—200 тыс. лет, от неандертальцев среднего палеолита – 200—40 тыс. лет, а современный человек появился 40—13 тыс. лет назад (поздний палеолит); от 13 до 5 тыс. лет отделяют нас от мезолита и неолита и примерно 5 тыс. лет длится историческая эра. Одно поколение длится около 25 лет, и мы отделены от нашего звероподобного предка всего десятком тысяч поколений.
Что мог за это время сделать естественный отбор?
3.1. Понятие о каналах эволюции и об ортогенезе
Эволюция вида одновременно идет в разных направлениях, но с очень разной скоростью. Молекула гемоглобина человека, отделившегося от своего общего предка с гориллой несколько миллионов лет назад, отличается от молекулы гемоглобина горилл лишь одной кислотой из 147, входящих в состав бета-цепи гемоглобина. Но в десятки раз большее время потребовалось, чтобы из тапирообразной морды вырос хобот слона и сформировалась шея жирафа, открывшая виду, устремившемуся по пути ее удлинения, массу листвы, недоступной другим нелазающим млекопитающим. Эволюция вида идет направленно, по определенному видовому каналу.
В сохранении жизни различных подвидов и пород тутового шелкопряда решающее значение приобрела регуляция числа поколений (вольтинности), развивающихся в течение года, так как в отсутствие этой регуляции вылупляющиеся гусеницы оказывались без корма, а выходящие из кокона бабочки откладывали яйца, обреченные на гибель; с другой стороны, в условиях тропиков лишнее четвертое, пятое, шестое, седьмое, восьмое поколение давало возможность решающим образом обогнать по темпам размножения подвид или породы, дающие меньшее число поколений. Естественно, что вольтинность «обросла» множеством корреляций, например, с темпами эмбрионального и гусеничного развития, с числом и размером откладываемых яиц, размерами тела, с устойчивостью к условиям температуры и влажности, со сложной системой нейрогуморальной регуляции, стрелочно направляющей развитие следующего поколения в разных направлениях – в направлении ли перерыва развития на эмбриональной стадии, или в направлении ее завершения и выхода гусениц.
Однажды создавшаяся система корреляции определяет и основные направления, «каналы», «туннели», движением по которым вид будет в дальнейшем реагировать на отбор.
Тутовый шелкопряд под влиянием отбора, направленного по каналу его эволюции, способен за десяток поколений пройти путь наследственного сдвига от огромной бабочки с коконом, весящим 3 г, до карликовых «подвидов» с весом в 6—7 раз меньшим (но одновременно, и, главное, дающих вместо одного поколения за год три, четыре, пять поколений).
Возникает лишенный всякой мистики ортогенез, направленность эволюции, и Вавилонским гомологическим рядам наследственной изменчивости начинают соответствовать гомологические корреляционные сдвиги под действием отбора.
Широко известно, например, непрерывное удлинение конечностей от гиппариона к лошади, ортогенез, связанный с господствующим путем приспособления – увеличением быстроты бега.
Прослеживая эволюцию и вымирание ископаемых ящеров, можно заметить, что самое мощное оружие нападения не спасло бы его обладателя от поражения в схватке с менее вооруженным хищником, если бы рука об руку со средствами нападения не существовало достаточно прочной защитной брони. Широко известны и коррелированные сдвиги в направлении повышения мощности средств защиты и нападения, закончившиеся бесчисленными тупиками эволюции в виде гигантских ящеров.
Основной канал эволюции с сопутствующими корреляциями постоянно становился главным объектом отбора, этот канал воспринимал основное давление отбора, хотя, конечно, отбор шел всегда и по множеству некоррелированных признаков.
3.2. Каналы эволюции человека
Понимание того, что в ходе эволюции предков человека наиболее интенсивный отбор вел их по пути всевозрастающей социализации, в частности по пути создания анатомической базы для членораздельной речи, по пути увеличения мозга, особенно его лобных долей, и «коры руки», необычайно важно. Только так можно понять, каким образом создавались и развивались биологические основы социальных эмоций, как бы эти основы мы себе ни представляли: в форме ли некоторых безусловных рефлексов, или, например, нервных связей лобных, префронтальных височных долей с центрами удовольствия или боли, либо же в виде готовности мозга к образованию экстраполяционных рефлексов (Крушинский Л.В., 1960), либо других, пока только расшифровываемых материальных субстратов.
Одно из важнейших событий, происшедших в начале длинной стадии древесного обитания наших предков, заключалось в сильном укорочении выступающей части морды. Опознание и наблюдение окружающего постепенно переместилось с морды и носа, с обоняния – на глаза, руки, на рассматривание и хватание.
Чувство обоняния у собак, по самым осторожным подсчетам, в 1000 раз, если не в миллион и даже многие миллионы раз острее, чем у человека, и они способны обонять, по выражению Д. Морриса (Morris D., 1967) «целые ландшафты запахов».
У обезьяны площадь двигательной коры, управляющая рукой, примерно также велика, как и площадь коры, управляющая ступней, тогда как у человека «кора руки» гораздо более велика, чем «кора ступни» – прямой результат отбора на более ловкое использование орудий. Сравнение емкостей, внутренних поверхностей черепа и их функций у современного человека и его предков показывает, что различия заключаются прежде всего в сравнительных объемах лобных долей центров речи, центров памяти, центров координации движения рук и пальцев.
Когда наш предок начал ходить на задних лапах, а передние лапы стали руками, появились орудия, начал стремительно расти мозг (табл. 1), начал слагаться совершенно новый канал коррелированного, сверхбыстрого эволюционирования.
Таблица 1
Темпы увеличения емкости черепа в ходе эволюции гоминид
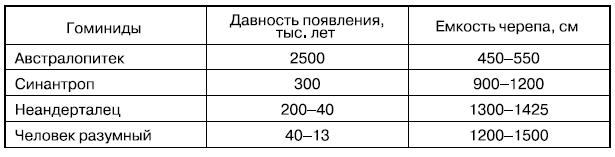
От австралопитека до неандертальца за 1—2 млн лет емкость черепа возросла втрое (Dobzhansky Th., 1956).
Заслуживает, впрочем, внимания и то, что от неандертальца до нашего времени емкость черепа не выросла: за счет других частей мозга вырос объем лобных долей.
Большой мозг беспомощен, пока его содержимое не связано в целое условными и экстраполяционными рефлексами, памятью, опытом, знаниями, умениями, навыками. Параллельно эволюционному росту мозга все более удлиняется срок беспомощности детенышей, срок, в течение которого они нуждаются в помощи и охране со стороны не только родителей, но и всей стаи, стада, орды, племени, семьи. Даже у самых примитивных племен ребенок до шести лет совершенно не способен к самостоятельному существованию, к обороне, и даже у индейцев лишь с девяти лет начинает самостоятельно охотиться. Непрерывная охрана, непрерывная кормежка беспомощных детей и беременных, численность которых составляла не меньше трети стаи, а иногда и большинство ее, могла осуществляться только стаей, ордой в целом, скованной в своей подвижности этой массой нуждающихся в охране и пище носителей и передатчиков генов. И если эволюция человека от питекантропа оставила заметные следы в виде постепенно меняющихся скелетов, то в отношении наследственных инстинктов и безусловных рефлексов человек должен был дальше отделиться от питекантропа, чем выводковые птицы от гнездовых.
Церебрализация направила становление человечества по еще более узкому каналу коррелированной эволюции из-за ряда дополнительных причин.
1. Хождение на задних конечностях изменило форму таза и лишило праженщин свойственной обезьянам способности рожать большеголовых детенышей. Поэтому подъем на задние конечности, появление прямостоящего питекантропа повлекли за собой рождение малоголовых детей с непрочным черепом, которому предстояло сильно и долго увеличиваться уже в ходе последующего развития, с незрелой нервной системой, тоже долго развивающейся после родов.