Эволюция кооперации и альтруизма: от бактерий до человека - Страница 9
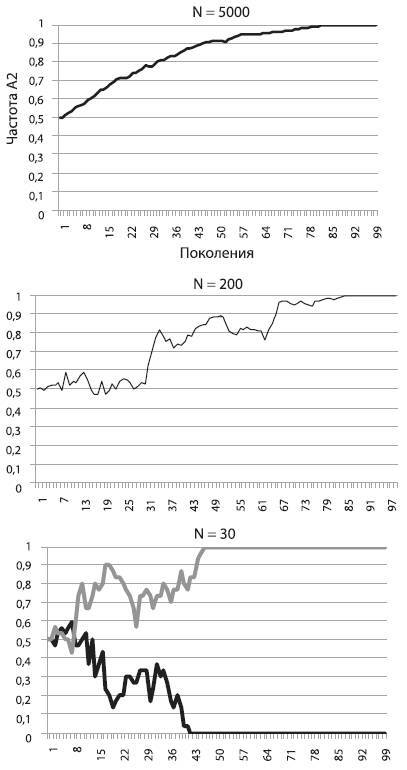
На среднем графике мы видим, какая судьба ждет такую же полезную мутацию, повышающую приспособленность на 5 %, в популяции меньшего размера (N = 200). Частота полезного аллеля и в этом случае росла и в конце концов достигла 1 (мутация зафиксировалась), но путь был труден и извилист. Пожалуй, нашей мутации могло и не повезти, ее шансы на фиксацию не были стопроцентными.
Наконец, на правом графике мы видим, что происходит с точно таким же полезным аллелем в совсем крошечной популяции (N = 30). На рисунке показаны результаты двух запусков модели. В одном случае мутация зафиксировалась, в другом – элиминировалась. Не правда ли, картинка похожа на результат работы дрейфа, а не отбора?
Так оно и есть. В этом заключается главный урок, который мы можем извлечь из наших экспериментов. Чем меньше популяция, тем слабее в ней власть отбора и тем могущественнее дрейф. В маленьких популяциях слабополезные и слабовредные мутации начинают вести себя фактически как нейтральные. Их частоты «случайно блуждают», пока не наткнутся на верхний или нижний порог. В маленьких популяциях, где царствует дрейф, небольшие различия в приспособленности становятся невидимыми для отбора. Поэтому слабовредная мутация запросто может зафиксироваться, а слабополезная – элиминироваться.
Хорошо это или плохо? В большинстве случаев, конечно, плохо. Потеря слабополезных мутаций мешает маленькой популяции приспосабливаться к меняющимся условиям. Бесконтрольное накопление слабовредных мутаций и вовсе может поставить ее на грань вымирания. Именно в этом, между прочим, видят биологи одну из причин того, что крупные животные вымирают в среднем чаще, чем мелкие. У крупных животных, таких как носороги или слоны, популяции не могут быть такими же большими, как у мышей или насекомых. Это снижает приспособляемость крупных животных.
Но у этой медали есть и обратная сторона. У маленьких популяций больше шансов выйти из «ловушки локального оптимума», т. е. сползти с невысокого пика ландшафта приспособленности и взобраться на другой, повыше. Ведь отбор гонит организмы вверх и только вверх. Если популяция велика и малейшее различие в приспособленности «заметно» для отбора, спуск по склонам становится невозможен. Однажды взобравшись на одинокий холм, большая популяция уже никогда с него не слезет. Что касается дрейфа, то он ведет организмы по ландшафту приспособленности хаотическим образом, не замечая подъемов и спусков. Если популяция невелика и дрейф силен, у организмов есть шанс иногда двигаться не только вверх, но и немного вниз (и в сторону). Спустившись в ложбинку, организмы могут «обнаружить», что отсюда есть другой, более перспективный подъем. Если, конечно, не вымрут раньше, чем на него наткнутся.
Вселенная древних белков продолжает расширяться
В 2010 году журнал Nature опубликовал интересную статью об эволюционном движении белков по ландшафтам приспособленности (Povolotskaya, Kondrashov, 2010). Авторы работы решили сравнить аминокислотные последовательности 572 древних белков, которые имелись уже у последнего общего предка всего живого[21] и были унаследованы его потомками, а также нуклеотидные последовательности соответствующих генов 836 прокариот (бактерий и архей). Эти белки называют древними, потому что после более чем 3,5 млрд лет эволюции в телах разнообразных потомков LUCA они до сих пор сохранили сходство своих аминокислотных последовательностей (вплоть до поразительного 40-процентного сходства рибосомных белков L14 у бактерий и архей) и продолжают выполнять те же функции, что и у Луки.

Расширение физической и белковой Вселенной. Стрела времени направлена сверху вниз. Слева: в ходе расширения физической Вселенной увеличиваются расстояния от произвольно выбранного объекта (например, Земли) до других объектов, причем скорость удаления пропорциональна расстоянию. Справа: в ходе расширения «белковой вселенной» дочерние молекулы удаляются в пространстве последовательностей от исходной точки, которая соответствует одному из белков LUCA. При этом дистанции между произвольно выбранным объектом (например, белком, принадлежащим эволюционной линии, которая привела к кишечной палочке E. coli) и другими объектами (родственными белками других эволюционных линий) постепенно растут. Однако белки с данной функцией не могут выйти за пределы «своей» подобласти в пространстве последовательностей – иначе это будет уже другой белок, выполняющий другую функцию (эволюционные события, связанные со сменой функции белка, в обсуждаемой работе не рассматриваются). Границы этой функциональной области показаны внутренним кругом. Рано или поздно расходящиеся белки достигнут этой границы, и дальнейшее расхождение станет невозможным. Стрелочками показано направление движения белков в пространстве последовательностей. По рисунку из Povolotskaya, Kondrashov, 2010.
Для начала Поволоцкая и Кондрашов решили выяснить, достигли уже эти белки максимального расхождения в пространстве последовательностей или их расхождение (накопление различий) все еще продолжается. Сохранение сходства после 3,5 млрд лет независимой эволюции, казалось бы, говорит о том, что возможности изменения этих белков ограниченны. Можно предположить, что области в пространстве последовательностей, соответствующие их функциям, невелики и предел расхождения давно достигнут. Но возможно и другое объяснение. Эти области могут быть велики, но труднопроходимы. Например, они могут представлять собой лабиринт из узких хребтов, разделенных пропастями, и поэтому на «освоение» всего доступного пространства не хватило миллиардов лет, прошедших со времен LUCA.
Исследователи проводят аналогию между эволюцией белков и расширением Вселенной. Эдвин Хаббл обнаружил, что галактики удаляются друг от друга, причем расстояние между галактиками положительно коррелирует со скоростью их разлетания. Экстраполируя эту тенденцию в прошлое, Хаббл пришел к выводу, что разлетание должно было начаться из одной точки. Эта идея легла в основу теории Большого взрыва. Нечто подобное происходит и с белками, расходящимися от общего предка – исходного белка с данной функцией, который был закодирован в геноме LUCA.

Принцип анализа белковых последовательностей, примененный Поволоцкой и Кондрашовым. Использовались четверки гомологичных белков, родственные отношения между которыми изображены в виде дерева. Анализ такой четверки позволяет понять, растет или снижается сходство между первыми двумя белками и четвертым. В данном случае считается, что у первого белка «предковая» (имеющаяся у белков 2 и 3) аминокислота R в 11-й позиции (выделена жирным шрифтом) заменилась на E, что привело к росту сходства первого белка с четвертым, т. е. к их сближению в пространстве последовательностей. У второго белка «предковая» аминокислота K в 3-й позиции (выделена полужирным шрифтом) заменилась на Y, что привело к снижению сходства второго белка с четвертым, т. е. к их расхождению в пространстве последовательностей. По рисунку из Povolotskaya, Kondrashov, 2010.
Чтобы выяснить, закончилось ли уже расширение «вселенной древних белков» или оно продолжается до сих пор, авторы применили оригинальные методы анализа белковых последовательностей. Нам будет полезно с ними ознакомиться – хотя бы для того, чтобы понять логику биоинформатики, молодой науки, занимающейся сравнением и осмыслением генетических текстов.
Использовались четверки гомологичных (происходящих от одного предка) и выполняющих одну функцию белков. Первые два белка в каждой четверке – это близкородственные молекулы с похожими аминокислотными последовательностями. Эти два белка назывались «сестринскими», и именно их эволюция анализировалась в рамках данной четверки.