Центральная нервная система. Анатомия и физиология - Страница 7
На рис. 27 в качестве активного обозначен тот перехват, в котором в текущий момент времени формируется потенциал действия. Между этим и соседними (невозбужденными) перехватами возникают силовые линии. Это должно привести к открытию в них натриевых каналов и создать условия для формирования потенциала действия. Но это произойдет лишь в одном из них – более удаленном от тела клетки. Другой же перехват находится в это время в состоянии невозбудимости, поскольку в нем как раз завершилось формирование потенциала действия и работает натрий-калиевый насос. Это состояние невозбудимости называется рефрактерным периодом. То есть вслед за потенциалом действия по аксону перемещается рефракторность, которая препятствует обратному распространению возбуждения.
Сальтаторное проведение имеет преимущество перед обычным способом проведения возбуждения по мембране, не покрытой миелином. Во-первых, этот способ более экономичен. Расстояние между перехватами Ранвье может достигать 2 мм, а ширина самого перехвата, как правило, не превышает 14 мкм. Поэтому энергозатратные насосы работают на площади, составляющей менее 1 % мембраны аксона. Еще одно преимущество сальтаторного проведения – его скорость, которая может достигать 120 м/с.
Нейроны контактируют между собой с помощью особых образований, называемых синапсами (рис. 28).
Основными компонентами синапса являются: пресинаптическая мембрана (мембрана передающего нейрона), синаптическая щель (пространство шириной 20–50 нм, заполненное межклеточной жидкостью) и постсинаптическая мембрана (мембрана принимающего нейрона).
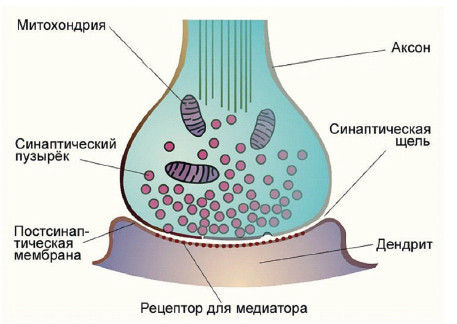
Рис. 28. Синапс
Способ передачи сигнала в подавляющем большинстве синапсов – химический. В синаптических пузырьках содержится особое вещество – медиатор. Когда потенциал действия приходит к окончанию аксона, деполяризованная пресинаптическая мембрана притягивает и разрывает пузырьки. Их содержимое выбрасывается в синаптическую щель, медиатор диффундирует к постсинаптической мембране и взаимодействует с ее рецепторными белками. В результате в постсинаптической мембране открываются определенные ионные каналы, а возникающий ионный ток меняет ее заряд, формируя постсинаптический потенциал. Чем выше частота потенциалов действия, тем сильнее воздействие на принимающий нейрон. Один потенциал действия вызывает выброс 200–300 квантов медиатора. Один квант открывает порядка 2 тысяч ионных каналов в постсинаптической мембране. Весь процесс синаптической передачи занимает около 2 мс. Таким образом, проведение в синапсах всегда одностороннее и осуществляется с временной задержкой.
Все синапсы в нервной системе делятся на возбудительные и тормозные (рис. 29).

Рис. 29. Возбудительные (В) и тормозные (Т) синапсы
Стрелками показано направление перемещения ионов через постсинаптическую мембрану; графики отображают процесс формирования возбудительного и тормозного постсинаптических потенциалов.
Возбудительные деполяризуют постсинаптическую мембрану (снижают разницу заряда внутренней и внешней поверхности мембраны) и формируют возбудительный постсинаптический потенциал (ВПСП). Тормозные синапсы гиперполяризуют мембрану принимающего нейрона (увеличивают разницу заряда) и формируют тормозной постсинаптический потенциал (ТПСП). Таким образом, возбудительные синапсы возбуждают принимающий нейрон, а тормозные – тормозят. Тип синапса определяется тем химическим веществом (медиатором), которое выбрасывается в синаптическую щель и воздействует на постсинаптическую мембрану.
Постсинаптические потенциалы по своим свойствам отличны от потенциалов действия. Потенциалы действия имеют постоянную амплитуду, распространяются без затухания и не взаимодействуют. Постсинаптические потенциалы, напротив, градуальны (имеют переменную амплитуду), локальны (распространяются по мембране с затуханием) и способны к суммации.
В норме нейроны постоянно испытывают множество синаптических воздействий. ВПСП и ТПСП «стекают» по мембране клетки в направлении аксонного холмика. Поскольку распространение происходит с затуханием, чем ближе синапс располагается к триггерной зоне, тем выше его «вес». Встречаясь по пути следования, постсинаптические потенциалы суммируются: одноименные усиливают друг друга, противоположные ослабляют. Если в какой-то момент времени суммарный ВПСП в области аксонного холмика превысит суммарный ТПСП на определенную (пороговую) величину, произойдет формирование потенциала действия.
1. Что отличает нервную клетку от других клеток организма?
2. Какова функция дендритов?
3. Каковы основные функции мембраны нейрона?
4. Какие факторы определяют проницаемость мембраны нейрона?
5. Какова природа мембранного потенциала покоя?
6. С каким мембранным процессом связана деполяризация мембраны?
7. С каким мембранным процессом связана реполяризация мембраны (возвращение заряда к исходному уровню)?
8. Что происходит во время работы ионного насоса?
9. Каковы особенности проведения в синапсах?
10. Каковы свойства постсинаптических потенциалов?
1. Что представляет собой серое и белое вещество ЦНС?
2. Что представляет собой псевдоуниполярный нейрон?
3. Каковы особенности организации узловой нервной системы?
4. Что отличает нервную клетку от других клеток организма?
5. Каковы структурные элементы нервной клетки?
6. Что в нервной системе является единицей информации?
7. В чем отличие безусловных и условных рефлексов?
8. В чем проявляется сигнальное значение условного рефлекса?
9. Как безусловные рефлексы классифицируются по уровню сложности?
10. Что такое доминанта, и каково ее значение?
11. Какие факторы определяют проницаемость мембраны нейрона?
12. Какова природа мембранного потенциала (потенциала покоя)?
13. Какие процессы реализуются на мембране нейрона после того, как деполяризация достигает пороговой величины?
14. В чем особенность сальтаторного проведения?
15. Опишите механизм синаптической передачи.
Глава 2
Организация спинного мозга и каудальных отделов ствола
Мы переходим к рассмотрению вопросов, касающихся строения и функционирования основных отделов ЦНС, выделяемых по анатомическим признакам. В этой главе речь пойдет о спинном мозге и каудальных отделах ствола головного мозга: продолговатом мозге, Варолиевом мосте и среднем мозге.
Спинной мозг – самая древняя часть ЦНС и самая просто организованная. Каудальные структуры ствола являются результатом эволюционного развития верхнего отдела спинного мозга. Их появление обусловлено необходимостью наращивания и совершенствования спинальных функций.
В филогенезе формирование ствола начинается с утолщения верхнего отдела спинного мозга и его «разворачивания» с дорсальной стороны, в результате чего его правая и левая половинки расходятся в разные стороны. Центральный канал расширяется, образуя четвертый мозговой желудочек. Серое вещество, представляющее собой в спинном мозге единую структуру, разделяется на ядра. Афферентные нейроны, находящиеся в спинном мозге в дорсальной части серого вещества, формируют чувствительные (сенсорные) ядра, которые в продолговатом мозге оказываются расположенными ближе к боковой поверхности. Эфферентные нейроны, находящиеся в вентральной части серого вещества, образуют моторные ядра, которые располагаются ближе к средней линии. Вегетативные ядра занимают промежуточное положение.
В спинном мозге связь с органами и тканями осуществляется с помощью спинномозговых нервов, в стволе эволюционным развитием этого инструмента становятся черепномозговые нервы.